

Наукова робота. Фізіолого-біохімічні зміні в дозріваючому насінні видів роду Acer L. в умовах урбоценозу
Роботу виконала:
Кіжнер Маріанна Ігорівна,
учениця 10 класу
КЗО НВО № 28
Дніпровської міської ради
Науковий керівник:
Богуславська Людмила Володимирівна,
вчителька біології
КЗО НВО№ 28
Дніпровської міської ради,
лауреатка IV Всеукраїнського Інтернет-конкурсу
„УЧИТЕЛЬ РОКУ–2019” за версією
науково-популярного природничого журналу
„КОЛОСОК” у номінації „Біологія”
ВСТУП
Важливе місце в системі заходів у нашій країні займають питання підвищення продуктивності лісів та інтенсифікація зеленого будівництва в містах і селищах. Серед деревних порід, яким належить зайняти чільне місце в цих заходах, одними з перших слід назвати клени. Велика кількість видів і садових форм, відмінні декоративні якості дерев, досить швидкий ріст, грунтополіпшуюча роль, високі фізико-механічні показники деревини – це ставить клени в один ряд з найціннішими деревними породами, що використовуються у народному господарстві. Їх широко використовують у зеленому будівництві для створення алей, груп і зелених масивів, а в лісовому господарстві–для створення високопродуктивних насаджень. Зелені насадження – невід’ємна складова навколишнього природного середовища та, зокрема, міських екосистем. Вони виконують важливі екологічні функції, до яких належать: очищення та збагачення повітря киснем, водоохоронні, водорегулюючі, ґрунтотзахисні, санітарно-гігієнічні функції. Позитивний вплив лісових насаджень проявляється в збереженні ґрунту від водної та вітрової ерозії, вони сприяють зменшенню поверхневого стоку, підтриманню рівня водності рік, запобігають замулюванню їх продуктами ерозії, поліпшують якість води, впливають на підвищення врожайності сільськогосподарських культур [1, 4].
Однією з важливих екологічних проблем в місті є проблема зелених масивів (міських парків, лісів, садів) [5]. Рослинність забезпечує комфортність умов проживання людей в місті, регулює газовий склад повітря і ступінь його забруднення. Озеленення вулиць займає особливе значення в покращенні екологічного стану міста, активно впливаючи на архітектурний вигляд та забезпечуючи в літню пору необхідний тіньовий режим пішіходам. В індустріальних центрах, де створюється найбільша загроза санітарному стану повітряного басейну, для оздоровлення міського середовища в околицях заводів рекомендується висаджувати клени американські [17, 18]. Вони є стійкими по відношенню до техногенного забруднення довкілля міста. Листя кленів поглинає значну кількість сірки. Також вони володіють антибактеріальною дією, позитивно впливаючи на стан повітряного середовища міста. Значне місце вони можуть зайняти в лісовому господарстві, особливо в спеціалізованих господарствах, через цінну деревину. Клени є акумуляторами пилу. Навіть і в зимові місяці, коли дерева позбавлені листя, вони мають велике пилозахисне значення [2, 12].
Кліматичні і грунтові умови ряду районів України дозволяють культивувати майже всі види кленів світової флори. При належній постановці інтродукційної роботи можна відібрати для збагачення дендрофлори України велику кількість дуже цінних видів клена.
Мета роботи – аналіз міжвидової мінливості роду Acer L. за комплексом фізіолого-біохімічних ознак. Для досягнення цієї мети були поставлені наступні задачі:
– провести оцінку еколого-біологічних властивостей колекції аборигенних та інтродукованих видів роду Acer L. ботанічного саду ДНУ;
– провести порівняльний аналіз вмісту легкорозчинних дозріваючого насіння видів роду Acer L.;
– визначити активність та ізоферментний склад пероксидази в дозріваючому насінні різних видів роду Acer L.;
– дослідити залежність ізоферментного складу антиоксидантів від рівня стійкості кленів до стресових умов.
Роботу виконано на кафедрі біохімії та біофізики факультету біології, екології та медицини Дніпропетровського національного університету імені Олеся Гончара.
РОЗДІЛ І ОГЛЯД ЛІТЕРАТУРИ
1.1 Походження і природний ареал окремих видів роду Acer L.
Клен (Acer L.) — рід дерев та кущів родини сапіндових (Sapindaceae). Існує біля 125 видів і 40 підвидів, батьківщина більшості з яких — Азія, але кілька видів також зустрічаються в Європі, північній Африці і Північній Америці. З початку ХІХ століття із заснуванням на Україні перших ботанічних садів розпочалася інтродукція іноземних видів клена. Зараз на Україні культивують близько 50 видів клена, причому багато з них стали досить відомими, а Клен ясенелистий (Acer negundo L.) і Клен цукристий (Acer saccaharum M.) настільки поширилися, що стали майже обов’язковими компонентами культурних насаджень.
Раніше клен класифікували до окремої родини кленових (Aceraceae), але зараз вважають частиною родини сапіндових. Клени широко поширені в Північній півкулі, від полярних областей Європи і Північної Америки до тропічних районів Центральної Америки і Південної Азії. В основному поширені в помірних широтах, в тропіках відомі лише кілька видів, а в Південній півкулі лише один вид – Клен лавровий (Acer laurinum L.), який доходить до острова Тимор в Індонезії. На території Африки клени присутні тільки на самій півночі, вздовж узбережжя Середземного моря, а в Південній Америці і Австралії відсутні зовсім. Клени в південній частині ареалу в основному віддають перевагу гірським районам, піднімаючись у висоту до 3000 м над рівнем моря в Гімалаях. На рівнинах зростає відносно невелика кількість видів. Ростуть ці дерева поодинці або невеликими групами, самостійні ліси утворюють вкрай рідко [3].
1.1.2 Еколого-біологічна характеристика роду Acer L.
Розкриття еколого-біологічних особливостей рослин неможливо без вивчення їх сезонного росту і розвитку. Вивченню сезонного розвитку рослин приділяється велика увага. При цьому під розвитком рослин розуміються «якісні зміни структури і функцій рослини і його окремих частин: органів, тканин і клітин, що виникають у процесі онтогенезу».
В.Н. Ворошилов (1960) прийшов до висновку, що сезонний ритм рослин правомірно розглядати як пристосувальну властивість, що виникає в процесі еволюції і закріплюються у генотипі для забезпечення оптимального рівня спряженості в часі процесів росту і розвитку з сезонними змінами цілого ряду факторів середовища проживання.
Численні дослідження свідчать про те, що температура повітря, особливо у високих широтах, є основним чинником, що безпосередньо впливають на пускові механізми, що здійснюють регуляцію ростових процесів. Початок лінійного росту пагонів слабо корелює з сумою позитивних температур. Як правило, в роки з швидким переходом середньодобової від негативних величин до позитивних, зростання пагонів починається при порівняно невеликій сумі позитивних температур, а в роки затяжної весни – при найбільшій їх сумі. Надалі інтенсивність росту пагонів також пов’язана з температурою повітря. Як правило, при значному підвищенні температури відзначається і більш інтенсивний ріст пагонів. Наступ кульмінації приросту пагонів теж пов’язано з температурою повітря. Він припадає на період з найбільш високою температурою повітря за ці місяці. Після кульмінації приросту відзначається слабкий зв’язок інтенсивності росту пагонів з температурою повітря. У цей період вона ще залишається порівняно високою, а приріст знижується досить швидко. Припинення росту пагонів незначною мірою залежить від температурного режиму.
Опади служать основним джерелом накопичення запасів ґрунтової вологи. Дослідження показали, що кількість і частота випадання опадів не позначаються на темпах зростання вегетативних органів дерева. Опади роблять помітний вплив на строки припинення діяльності камбію стовбура.
При цьому характер їх впливу на цей процес розрізняється у зв’язку з умовами зростання. В умовах надмірного зволоження ґрунтового формування деревини триває Тим довше, чим менше випадає опадів в період загасання зростання, і, навпаки, велика кількість опадів призводить до більш раннього припинення росту. Абсолютно протилежний ефект в умовах помірної зволоженості ґрунту.
Сонячне світло є джерелом енергії для фотосинтезу, ефективність якого визначається інтенсивністю сонячної радіації, яка може посилювати або сповільнювати цей процес. Тривалість і інтенсивність сонячного сяйва впливає на розвиток рослин як через зміну швидкості продукування асимілятів, так і через зміну температурного режиму [6].
Встановлено, що клен є породою достатньо стійкою до дії промислового забруднення. Відносний життєвий стан насаджень кленів в умовах промислових центрів оцінюється як «здорове». Пігментний комплекс кленів характеризується підвищеною стійкістю до збільшення рівня промислового навантаження, що проявляється у збільшенні вмісту хлорофілу в умовах забруднення [22].
Види роду клен широко використовуються в різних галузях народного господарства, особливо в озелененні. Деревина клена щільна, тверда і міцна, високо цінується в меблевій промисловості, столярній справі. Листи містять каучук, а кора дубильні речовини [20]. Життєві форми кленів – чагарник або дерево. Характерні риси – осіннє забарвлення крони, що варіюється від яскраво-жовтого до яскраво-пурпурового кольору. Представники родини кленових мають деревоподібний зовнішній вигляд і, як правило, не досягають великих розмірів. Клен характеризується рясним розгалуженням. З кожного вузла ростуть дві повністю супротивні гілки. Листя клена розділене на кілька часток зубчастого виду. Листи клена змінюють колір перед листопадом, у такий спосіб персоніфікуючи прихід осені. Плоди клена – крилаті, мають незвичайний і яскравий вигляд, складаються з двох однакових частин і при падінні обертаються, несучи насіння на значну відстань. Плід дозріває в проміжку від двох до шести тижнів після цвітіння [20].
Високо цінуються клени за свої декоративні якості: колоноподібні стовбури, розлогі, тінисті крони, красиве листя. Значна частина кленів є прекрасними медоносами. Клен польовий, татарський, гостролистий та ясенелистий рекомендовані як супутні породи полезахисних лісових смуг. Клени вітростійкі, швидко ростуть. Деякі види кленів мають значні шумозахисні властивості. До них відносяться клен сріблястий, польовий, татарський, до того ж вони мають досить високі фітонцидні властивості. Ряд видів виділяється ароматними квітами, а деякі – декоративними плодами [21].
1.2 Стійкість кленів до абіотичних факторів середовища
Більшість видів кленів у районах природного зростання приурочені до умов помірного та досить вологого клімату. Такі ж екологічні вимоги стосовно зволоженості у певній мірі притаманні і аборигенним видам кленів. Тому основним фактором, який обмежує можливості культивування у степовій зоні багатьох видів клена, є посушливі умови вегетаційного періоду.
Витривалість рослин в умовах гідротермічного стресу обумовлюється функціонуванням пристосувальних механізмів на різних рівнях структурної організації та життєдіяльності рослинного організму. У регулюванні водообміну рослин значна роль належить величині водоутримуючих сил та фракціонуванню внутрішньоклітинної води. Ці показники визначають швидкість водовіддачі і зрештою відображують стійкість клітин до зневоднення. Водоутримуюча здатність залежить від структурованості протопласта та вмісту у ньому гідрофільних колоїдів. У стійких рослин під час посухи спрямованість метаболічних процесів зсувається в бік підвищення гідрофільності протопласта, що пов’язано із зниженням водного потенціалу та збільшенням фракції міцно зв’язаної води [23]. У більшості видів кленів в умовах гідротермічного стресу підвищується вміст зв’язаної води, що відбувається за рахунок зменшення загальної оводненості листя і фракції лабільної води. Таку спрямованість перерозподілу фракційного складу води можна розглядати як адаптивну реакцію рослин на посуху [24].
Клени є посухостійкими рослинами, що характеризуються дещо меншою оводненістю тканин, ніж у малостійких рослин. Для підтримки стабільного функціонування посухостійким рослинам необхідна менша порівняно з мезофітними видами вологозабезпеченість.
Інтродуценти роду Клен в міських насадженнях мають виражені відмінності за рівнем жаростійкості листя, а також показником водного режиму листя та стабільності цих показників в різні роки вегетації. Найбільша жаростійкість була відмічена у листя кленів гостролистого, татарського,а також польового, найменша – у листя клену ясенелистого. Найбільша оводненість виявлена у листя клену ясенелистого,а найменша – у кленів татарського, гостролистого та польового. Показано, що у кленів гостролистого, татарського і польового водозатримна здатність листя у порівнянні з показниками загальної оводненості тканин досить висока [7].
Більшість видів роду Клен морозостійкі і не потребують додаткового укриття на зиму. Багатьом культурам достатньо висадки у захищені від вітру місця. Кореневу шийку молодих саджанців клена у суворі зими слід прикривати сухим листям. При пошкодженні морозом однорічних пагонів клену їх необхідно обрізати. Крона швидко відновлюється за рахунок нових пагонів. З віком морозостійкість підвищується. Вивчення морозостійкості кленів показали що у всіх видів спостерігається загальна тенденція до збільшення ступеню пошкоджень по мірі зниження температури і часу її впливу; для всіх органів рослин згубне різке падіння температури. Сильно знижується морозостійкість в результаті різких перепадів температури від більш низьких показників до вищих. Найвищою зимостійкістю характеризуються клени з раннім початком і короткою або середньою тривалістю вегетації, початок і завершення якої синхронізоване з динамікою фенологічних пор року.
Клен гостролистий досить морозостійкий. Дикі види є більш морозостійкими, ніж виведені культурні сорти. Клен ясенелистий витримує до -400С, однак морозостійкість значно коливається в залежності від віку рослини та походження насіння, з якого він вирощений [8].
Однією із найскладніших проблем підвищення лісистості степової зони є значне розповсюдження у її межах засолених ґрунтів. Загальна площа їх в Україні становить близько 4 млн. га. До засолених відносять ґрунти, які містять легкорозчинні солі, токсичні для рослин. Залежно до кількості та глибини залягання солей виділяють солончаки, солончакові та солончакуваті
Види ґрунтів, які виявляють негативну дію легкорозчинних солей на деревні та чагарникові породи. Окрім солей негативним у засолених ґрунтах є наявність поглиненого натрію (Nа+), який зумовлює дуже негативні водно-фізичні властивості ґрунтів: в’язкість, набувняніння у вологому стані, щільність у сухому. Виділяють різного ступеню солонцюваті ґрунти (Nа+ 5–20%) та солонці (Nа+ >20%). Несприятливі властивості засолених ґрунтів виявляються в умовах недостатньої вологозабезпеченості. Токсичність легкорозчинних солей та незадовільні водно-фізичні властивості засолених ґрунтів зумовлюють їхню надто низьку лісопридатність. Кращими лісорослинними властивостями вирізняються ґрунти зі слабкою засоленістю, на яких при доволі високому рівні агротехніки можливе створення відносно стійких насаджень із солевитривалих деревних і чагарникових порід. На ґрунтах вищого ступеня засолення (середньо – і дуже засолених) створення деревних насаджень можливе лише після проведення корінних меліорацій.
Клени – солестійкі рослини,не вимогливі до ґрунтів. Серед багатьох видів кленів солестійкими є клен польовий і клен татарський. Слабосолестійкими – клен ясенелистий, гостролистий та сріблястий [11].
Клени акумулюють важкі метали у надлишкових кількостях. Дослідження кленів гостролистого та ясенелистого показують,що вміст свинцю в насінні клена гостролистого знаходився на рівні 1.90, а в клені ясенелистому 1.10 мг/кг, а кадмію коливався у обох видів у межах 0.15-
0.26 мг/кг сухої маси. Вміст нікелю у досліджуваних видів кленів були рівні – 4.4–6.1 мг/кг. Найвищі показники вмісту в насінні зареєстровані для цинку: 45.4 для A. platanoides L. і 68.9 мг/кг для A. negundo L. [16].
При порівнянні величин вмісту важких металів у насінні аборигенного виду A. platanoides L. в умовах міста і в сільській місцевості виявлено, що вміст кадмію, нікелю і цинку був більшим в умовах міста, ніж в сільській місцевості на 18, 39 і 42% відповідно. Рівень кадмію в насінні обох моніторингових ділянок був практично однаковим. Інша картина спостерігалась для насіння інтродукованого виду A. negundo L. Так, встановлено, що вміст свинцю, кадмію і нікелю був зниженим у насінні цього виду на забрудненій викидами автотранспорту території порівняно з сільською місцевістю на 19,7 і 22% відповідно. Рівень цинку в насінні клена ясенелистого був практично однаковим для досліджуваних територій [14].
Наслідком забруднення навколишнього середовища викидами автотранспорту виявилась зміна маси 1000 насінин. Так, спостерігається зниження цього показника у клена гостролистого на 13% у порівнянні з контролем. Маса 1000 насінин, зібраного в сільській місцевості, становила 65.6 г, а насіння, зібраного уздовж автомагістралі – 57.3 г. У клена ясенелистого зниження маси дослідного насіння було на рівні 10% у порівнянні з контролем. Зниження маси насіння пов’язано з погіршенням його якості, так як збільшується кількість порожніх насінин. Найвищу здатність до накопичення в насінні двох видів кленів виявив цинк, потім нікель, свинець, а кадмій мав найменшу здатність до поглинання. Виявлено видову специфічність у накопиченні важких металів в насінні видів роду Acer L. [9].
Посилення забруднення середовища, особливо у великих містах, збільшення розмаху будівництва доріг і водойм свідчать про зростання актуальності озеленювальних робіт.
Стійкість до загазованості і задимлення, висока декоративність, невибагливість висувають багато видів Aceraceae на перше місце серед перспективних деревних інтродуцентів. Відомо, що місцеві деревні породи часто не можуть успішно протистояти отруйним промисловим викидам. Багато видів роду Acer L. мають в цьому плані ряд переваг, так як мають підвищену стійкість до задимлення та загазованості повітря, високу декоративність і здатність виділяти фітонциди.
Газостійкість деревних порід залежить від декількох чинників і внутрішньо-біологічних особливостей виду, комплексу ґрунтово-кліматичних умов, температури та вологості повітря, віку рослин, пори року. Впливу шкідливих речовин повітря зазнають насамперед молоді рослини,їх листя і пагони. В оптимальних ґрунтово-кліматичних умовах газостійкість завжди вища. З підвищенням температури повітря газостійкість рослин знижується. З підвищенням вологості повітря газостійкість також зменшується, оскільки частина газів розчиняється у краплинах води, що сприяє потраплянню на листкову поверхню більшої кількості шкідливих речовин. Порівняно стійким до шкідливих речовин вважається клен польовий. Слабостійкими є клен ясенелистий і татарський. Клен гостролистий не є стійким [15].
У механізмах стiйкостi i адаптації рослин кленів до промислових забруднювачів суттєву роль відіграють запасні білки як інтегральні показники перебігу фізіолого-біохімічних процесів у репродуктивних органах. В забрудненій зоні спостерігається зниження кількості білку, що свідчить про значний вплив інгредієнтів викидів автотранспорту на фізіологічні та метаболічні процеси, як всієї рослини, так і насіння в процесі його формування [13].
В результаті проникнення газів у рослинні клітини, в них накопичуються шкідливі речовини, які порушують процеси обміну речовин. Більш газостійкі рослини мають добре розвинену покривну тканину листя і міцну структуру палісадної та губчатої паренхіми [25].
РОЗДІЛ ІІ ОБ’ЄКТ, МАТЕРІАЛИ ТА МЕТОДИ ДОСЛІДЖЕННЯ
2.1 Природно-кліматичні умови Центрального Степу України
Клімат Центрального Степу помірноконтинентальний. Сумарна сонячна радіація характеризується найвищими в Україні показниками. Для Степу властива й найвища багаторічна величина радіаційного балансу. Пересічна температура липня становить +20…+24°С, січня -2…-9°С. Вегетаційний період триває 210-245 днів. Зона піддається значному впливу циклонів особливо у зимовий період. Циклони здебільшого надходять з півдня і південного заходу — на північ і північний схід [23].
Зазвичай за рік у Степу випадає 300-450 мм опадів. Часто бувають посухи. Значної шкоди сільському господарству завдають суховії, пилові бурі, особливо навесні та в ранньолітні періоди вегетації. Загалом кліматичні та агрокліматичні ресурси зони сприятливі для ведення сільського господарства, особливо в разі зрошення посушливих земель. Серед рослинного покриву переважає типова степова трав’яна рослинність: різнотравно-ковилово-типчакові степи (степові чагарники і дерева — терен, вишня). З середини літа степ вигорає. Деревна рослинність трапляється в глибоких долинах, на заплавах річок, чагарники – в ярах і балках [19].
2.2 Об’єкт дослідження
Об’єктами дослідження було обрано дозріваюче насіння двох видів роду Acer L. – клен гостролистий, рослина, яка є аборигенним видом, тобто зростає природно на території Дніпропетровської області, та клен ясенелистий – інтродукований вид, завезений до Європи із Північної Америки. Моніторинговими ділянками слугували зелені насадження кленів вздовж автомагістралі вздовж проспекту Богдана Хмельницького, де спостерігається інтенсивний потік автомобільного транспорту. Контрольні проби були взяті з дерев в с. Миколаївка Софіївського р-ну Дніпропетровської області (умовно чиста зона). Село розташоване на пологих схилах балочного ландшафту в долині річки Базавлук, в 100 км від міста Дніпропетровська.
Дослідження проводили в період дозрівання насіннєвого матеріалу. Було обрано по 3 дерева кожного виду в забрудненій та умовно чистій зонах, з яких потім проводилися відбори за наступними датами: 1 – 7-14.07.2017; 2 – 7-14.08.2017; 3 – 7-15.10.2017. З насіння зібраного в липні не вдалося провести досліди, так як воно було ще не повністю сформованим.
Насіння звільняли від зовнішньої оболонки, розтирали у фарфоровій ступці з пестиком до порошкоподібної суміші (попередньо залишивши по 1000 насінин кожного виду зібраного в жовтні для подальшого знаходження маси). Насіння зберігали в паперових пакетах в темноті на холоді.
Рослини визначалися за Шепотьєвим та Кохном [26, 27]. Назви рослин наведені за сучасною ботанічною номенклатурою [28].
2.3 Екстракція легкорозчинних білків із насіння кленів гостролистого та ясенелистого
Для вилучення білків наважку 0,1 г гомогенізованого матеріалу заливали 1 мл 0,1 М трис-НСІ буфера, рН 7,7 та екстрагували протягом 1,5 год при +40С, центрифугування проводили 15 хв при швидкості 15000 g при температурі +40С. Отриманий супернатант зливали у пробірки для визначення вмісту білку та визначення активності і складу пероксидази.
2.4 Визначення вмісту білка
Визначення вмісту білку проводили за методом Бредфорд [29], який базується на утворенні кольорових комплексів білку з розчином барвника (Кумасі діамантово-блакитного G–250).
2.5 Визначення активності пероксидази
Активність пероксидази визначали за методом Бояркіна [30].
Приготування ферментного препарату. Наважку рослинного матеріалу вагою 0,1 г розтирали у ступці з кварцевим піском і 1,0 мл ацетатного буферу з рН 5,4 до утворення однорідної маси. Отриманий гомогенат центрифугували на центрифузі К-24 протягом 20 хв. при 16000 g. Надосадкову рідину (супернатант) зливали в чисті пробірки і використовували для визначення активності пероксидази. Про активність пероксидази судили за часом утворення синього кольору окисленого бензидину. Для вимірювання активності брали таке розведення супернатанту, щоб зміна кольору відбувалась від 10 до 60 сек.
2.6 Дослідження ізоензимного складу пероксидази
Принцип методу заснований на специфічному прявленні пероксидази за допомогою гістохімічної кольорової реакції продуктів її ферментативного перетворення. Поліакриламідні гелі готують у вигляді пластин. 30 %-ний запасний розчин, який містіть 29,2 % (маса/об’єм) акриламіду і 0,8 % (маса/об’єм) N, N’ – метилен-біс-акриламіду, фільтрують і зберігають при 40С. Перед використанням розчин нагрівають до 200С і деаерують у вакуумі водоструйного насоса. Розділення розчинних білків проводять у 7,5 %-ному поліакриламідному гелі (рН 8,8) і 4,5 %-ному формуючому гелі (рН 6,8). Після внесення в електрофоретичну систему робочого гелю на поверхню гелю обережно нашаровують воду. По завершені полімеризації верхній шар вісмлктують, промивають буфером формуючого гелю. Перед цим у камеру вставляють гребінець для формування карманів. Залежно від об’єкта слід вносити в кожний карман від 50 до 100 мкг білка (точну кількість внесеного білка визначають експериментально). Густина досліджуваних екстрактів перед внесенням у кармани підвищується, якщо до них додати сахарозу (1 М розчину). Величина рН Трис-гліцинового буфера для електрофоретичної камери становить 8,3 Н. Буфер, який готується так: 6,0 г Трис-оксиметіламінометану, 28,8 г гліцину помістити в мірну колбу на 1 Л і довести до мітки; перед використанням цей розчин необхідно розбавити в 10 разів. У верхню камеру для електродного буфера додають декілька крапель 0,001 %-ного барвника бром фенолового синього. Тривалість електрофорезу становить 2,5-3,0 год при тем пера турі 40С і силі току 20 мА на гельову пластину.
Після проведення електрофорезу гелі фарбуються в інкубаційному середовищі:
а) запасний розчин: в 100 сл 7 %-ной розчинити 16 г та наситити розчин етилендіамінтетраоцтової кислоти (ЕДТА), потім профільтрувати. Фільтрат наситити бензидином (основою) і знову профільтрувати.
б) інкубаційне середовище: 9 мл запасного розчину та 1 мл 0,1 %-ного свіжовтготовленого розчину пероксиду водню, декілька кришталиків нітропрусиду натрію (10-20 мг).
Гелеві пластинки заливають інкубаційним розчином і додають кристали нітропрусиду. Кювету з пластинкою струшують, щоб кристали нітропрусиду розчинилися. Через 5-10 хвилин у зонах з пероксидазною активністю з’являється синє забарвлення. Далі інкубаційне середовище зливають і заливають гель водою [31].
Денситометруванння пластинок проводили за допомогою програми «Електрофор-менеджер», 2005 р.
2.7 Статистична обробка даних
Досліди виконані не менше як в 3-х повторностях. Отримані результати опрацьовували статистично за допомогою стандартних комп’ютерних програм у Excel 2003/XP. Достовірність різниці між варіантами оцінювали за критерієм Ст’юдента, використовуючи 5% рівень значущості [32].
РОЗДІЛ ІІІ РЕЗУЛЬТАТИ ДОСЛІДЖЕНЬ ТА ЇХ ОБГОВОРЕННЯ
Оцінка вмісту білку в дозріваючому насінні клена гостролистого та клена ясенелистого. Фізіолого-біохімічні методи аналізу розкривають механізми адаптації на молекулярному рівні, які є первинними відносно морфологічних змін. Внаслідок впливу внутрішніх та зовнішніх факторів у різні періоди життя материнської рослини, насіння як продукт її життєдіяльності також набуває різних змін, у тому числі і відхилення від норми метаболічних систем.
Білки є одним із головних чинників формування системи стійкості рослин до різних факторів середовища, в тому числі і при викидах автотранспорту. Є дані про білковий обмін вегетативних органів роду Acer L. у річному циклі розвитку [38]. Проте в літературі не вистачає інформації щодо вивчення накопичення запасного білку в процесі дозрівання насіння різних видів роду Acer L. Тому доцільно було дослідити зміну накопичення білків сім’ядолей насіння клена гостролистого та клена ясенелистого за умов викидів автотранспорту. Результати оцінки вмісту запасного білку в дозріваючому насіння різних видів роду Acer L., зростаючих в різних місцях (умовно чиста зона, біля автомагістралі) представлені в табл. 3.1, 3.2.
Таким чином, в насінні клена гостролистого, зібраному в чистій зоні спостерігається більший вміст запасного білка, ніж у рослин з забрудненої зони. Так, вміст запасного білку у насінні з с. Миколаївка становить 2,5 мг/мл у серпні, та 3,2 мг/мл у жовтні. Насіння, зібране вздовж автомагістралі по вул. Героїв Сталінграду має вміст білку відповідно у серпні – 2,1 мг/мл, у жовтні – 2,6 мг/мл. У порівнянні з контролем в насінинах із забрудненої зони виявлено зниження рівня білку: у серпні на 16%, у жовтні на 19%.
В насінні клена ясенелистого спостерігається в серпні зниження білку на 22%, тобто в чистій зоні вміст білку становить – 2,8 мг/мл, в забрудненій зоні – 2,2 мг/мл. У жовтні вміст білку, в насінні зібраного в с. Миколаївка становить – 3,3 мг/мл, а в насінні, зібраного вздовж автомагістралі – 2,7 мг/мл. Таким чином спостерігається зниження рівня білку на 18%.
Таблиця 3.1
Зміни вмісту запасного білку в дозріваючому насінні клена гостролистого за дії викидів автотранспорту
|
Місяць відбору |
Вміст білку, мг/мл |
Місяць відбору |
Вміст білку, мг/мл |
Відсоток до контролю |
|
|
Контроль |
Дослід |
||||
|
серпень |
2,5±0,02 |
серпень |
2,1±0,02 |
84 |
|
|
жовтень |
3,2±0,05 |
жовтень |
2,6±0,03 |
81 |
|
Примітка: похибка вибірки не перевищує 5% від середніх значень
Таблиця 3.2
Зміни вмісту запасного білку в дозріваючому насінні клена ясенелистого за дії викидів автотранспорту
|
Місяць відбору |
Вміст білку, мг/мл |
Місяць відбору |
Вміст білку, мг/мл |
Відсоток до контролю |
|
Контроль |
Дослід |
|||
|
серпень |
2,8±0,03 |
серпень |
2,2±0,01 |
78 |
|
жовтень |
3,3±0,04 |
жовтень |
2,7±0,03 |
82 |
Примітка: похибка вибірки не перевищує 5% від середніх значень
Отже, за отриманими даними можна вказати на те що в забрудненій зоні спостерігається зниження вмісту білку, що свідчить про значний вплив інгредієнтів викидів автотранспорту на фізіологічні та метаболічні процеси, як всієї рослини, так і на насіння в процесі дозрівання.
Дослідження зміни маси 1000 насінин в умовах забруднення викидами автотранспорту. Оцінка маси 1000 насінин використовується для фітоіндикації ступеню негативної дії забруднення середовища і є доцільним тест-параметром в моніторингових дослідженнях. В літературі зустрічаються дані щодо зміни маси 1000 насінин під впливом промислових емісій [34]. Проте значний інтерес становить, зміни маси насіння під впливом викидів автотранспорту. Тому вивчено, зміну маси зрілого насіння клена гостролистого та клена ясенелистого в чистій зоні, та забрудненій.
Таким чином, в умовах забрудненої зони маса 1000 насінин клена гостролистого знижується на 13 %, в умовах чистої зони маса становить – 65,6 г, а в зоні викидів автотранспорту – 57,3 г (рис. 3.1).
Для клена ясенелистого характерне зниження маси 1000 насінин на 10 % для дослідної зони у порівнянні з контролем. Маса 1000 насінин, зібраного в с. Миколаївка становить – 40,6 г, а в насінні зібраного вздовж автомагістралі становить – 36,7 г (рис. 3.1).
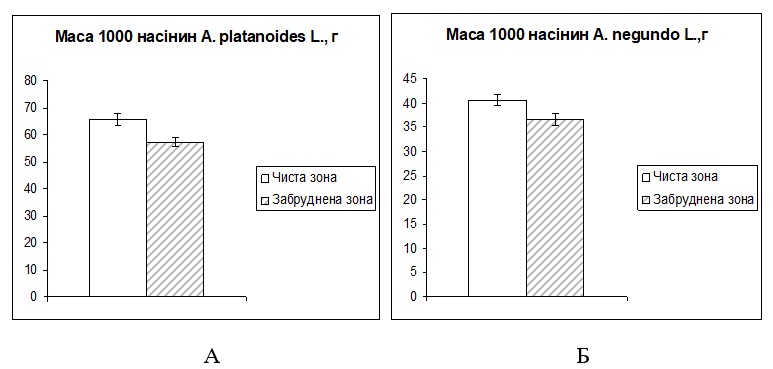
Рис. 3.1 – Зміна маси 1000 насінин A. platanoides L. (А) та A. negundo L. (Б)
Отже, зниження величини маси 1000 насінин свідчить про підвищення відсотка пустих насінин, або про зменшення розмірів насіннєвого матеріалу за умов викидів автотранспорту.
Активність пероксидази і ізоферментний склад в процесі дозрівання насіння. Для вивчення механізмів біохімічної адаптації деревних рослин до екстремальних умов техногенного забруднення середовища суттєве значення має дослідження фізико-хімічних властивостей ферментів. Важливу роль в захисних реакціях рослин на різноманітні стресові впливи належить пероксидазі.
Серед вивчених видів кленів показано різний рівень активності пероксидази, яка відіграє важливу роль у процесах дихання, росту і розвитку, захисних реакціях рослин. Доведено участь у захисних реакціях рослинних клітин бруньок ферменту пероксидази [35]. Пероксидазу вважають основною зимовою дихальною системою [36]. Деякими авторами підвищення стійкості до морозів пов’язується з підвищенням кількості ізоферментів пероксидази [37].
Здійснені дослідження показали зміни активності пероксидази за умов викидів автотранспорту. Зміни активності пероксидази в насінні клена гостролистого в процесі дозрівання представлені в табл. 3.11. Таким чином, спостерігаємо підвищення активності пероксидази в насінні клена гостролистого в серпні відповідно на 61 % і 87 %. В жовтні показники активності пероксидази в чистій зоні становили – 63,3, а в забрудненій зоні – 101,2. Тобто, в жовтні величина активності збільшується на 60 %.
Таблиця 3.11
Зміни активності пероксидази в процесі дозрівання насіння клена гостролистого (од.опт.густ./ г.хв.)
|
Місяць відбору |
Контроль |
Дослід |
% до контролю |
|
Серпень |
50,4±1,36 |
81,2 ±2,82 |
161 |
|
Жовтень |
63,3±2,20 |
101,2±1,12 |
160 |
Примітка: похибка вибірки не перевищує 5% від середніх значень
Результати дослідження зміни активності пероксидази в насінні клена ясенелистого наведені в табл. 3.12. В насінні, зібраного в серпні місяці в с. Миколаївка загальна активність пероксидази становить – 60, а в забрудненій зоні – 99,2. Тому, у порівнянні з контролем спостерігається збільшення активності на 65 %. У жовтні в порівнянні з серпнем активність пероксидази знижується. Проте у порівнянні з контролем також спостерігається підвищення активності на 59 %.
Отже, спостерігається значне підвищення активності пероксидази в дозріваючому насінні клена гостролистого та клена ясенелистого за умов викидів автотранспорту. Вважаємо, що це свідчить про те що клени, зростаючі у забрудненій зоні є сприйнятливими до дії викидів автотранспорту.
Таблиця 3. 12
Зміни активності пероксидази в процесі дозрівання насіння клена ясенелистого (од.опт.густ./ г.хв.)
|
Місяць відбору |
Контроль |
Дослід |
% до контролю |
|
Серпень |
60,0±2,82 |
99,2±3,51 |
165 |
|
Жовтень |
66,3±2,31 |
105,5±4,48 |
159 |
Примітка: похибка вибірки не перевищує 5% від середніх значень
Під впливом на рослини стресових факторів в клітинах формується унікальний стресовий набір ізопероксидаз. У рослин, не стійких до стресових впливів, генетичного потенціалу організму не вистачає для того, щоб сформувати оптимальний ізоферментий комплекс, і в критичних ситуаціях це може призвести до летального випадку. Пероксидази можуть нести достатню кількість інформації про фізіологічний стан рослин і можуть слугувати критерієм стійкості до стресових факторів.
Здійсненні дослідження показали зміни ізоферментного складу пероксидази у процесі дозрівання за дії викидів автотранспорту. Як видно, у процесі дозрівання насіння клена гостролистого як контрольних, так і дослідних зразків (табл. 3.13) присутні ізоформи з рІ від 3,55 до 7,20. Так, у контрольному варіанті присутні 8 ізоформ ферменту з рІ в діапазоні рН 3,65–6,90, у досліді спостерігається ізоформа з рІ 3,55, та ще 6 ізоформ з рІ в діапазоні рН 3,65–6,90 (серпень). Ізоформи з рІ 5,35 і 6,70 відсутні. У зрілому насінні клена гостролистого (дослід) виявлено 8 ізоформ ферменту з рІ від 3,60 до 7,20. У контролі відсутні ізоформи з рІ 3,60; 3,70;6,80 .
У процесі дозрівання насіння клена ясенелистого змінюється компонентний склад пероксидази (табл. 3.14). У насінні зібраному в серпні – контрольному зразку виявлено 8 ізоформ ферменту з рІ в діапазоні рН 3,60 – 6,45. У досліді 7 ізоформ з рІ 3,60 – 6,80, відсутні ізоформи з рІ 4,20; 4,22. Виявлено ізоформи з рІ 4,80; 6,80. У зрілому насінні (контроль) виявлено 7 ізоформ ферменту з рІ в діапазоні рН 3,65 – 5,25. У досліді присутні ізоформи з рІ 3,68; 6,10. Відсутня ізоформа ферменту з рІ 5,00.
Таблиця 3.13
Ізоферментний склад у процесі дозрівання насіння клена гостролистого
|
Значення рІ |
Серпень |
Жовтень |
||
|
контроль |
дослід |
контроль |
дослід |
|
|
3,55 |
– |
1,11 |
– |
– |
|
3,60 |
– |
– |
– |
3,34 |
|
3,62 |
– |
– |
28,95 |
– |
|
3,65 |
0,93 |
7,19 |
1,11 |
1,98 |
|
3,70 |
1,10 |
39,15 |
– |
1,77 |
|
3,71 |
– |
– |
0,24 |
11,00 |
|
4,65 |
22,62 |
4,65 |
– |
– |
|
5,20 |
8,01 |
5,70 |
– |
– |
|
5,35 |
0,36 |
– |
– |
– |
|
6,55 |
34,92 |
40,08 |
10,53 |
20,57 |
|
6,70 |
31,28 |
– |
– |
– |
|
6,80 |
– |
– |
– |
15,87 |
|
6,90 |
0,78 |
2,11 |
– |
– |
|
7,00 |
– |
– |
58,11 |
41,91 |
|
7,20 |
– |
– |
1,05 |
3,56 |
Таким чином, в дослідному насінні спостерігаємо зміни в ізоферментному складі пероксидазної системи. Виявлено в насінні за дії викидів автотранспорту наявність нових ізоформ. Це і обумовлює підвищення активності пероксидази в забрудненій зоні.
Таблиця 3.14
Ізоферментний склад у процесі дозрівання насіння клена ясенелистого
|
Значення рІ |
Серпень |
Жовтень |
||
|
контроль |
дослід |
контроль |
дослід |
|
|
3,60 |
1,92 |
3,91 |
– |
– |
|
3,65 |
2,78 |
2,25 |
0,72 |
2,49 |
|
3,68 |
– |
– |
– |
16,37 |
|
3,70 |
7,84 |
6,40 |
4,96 |
2,69 |
|
4,20 |
11,61 |
– |
– |
– |
|
4,22 |
1,23 |
– |
– |
– |
|
4,30 |
– |
– |
9,80 |
6,45 |
|
4,35 |
– |
– |
13,99 |
1,09 |
|
4,70 |
72,65 |
10,44 |
– |
– |
|
4,80 |
– |
7,80 |
– |
– |
|
4,85 |
1,07 |
– |
– |
– |
|
5,00 |
– |
– |
1,22 |
– |
|
5,10 |
– |
– |
67,00 |
4,86 |
|
5,25 |
– |
– |
2,30 |
59,81 |
|
6,10 |
– |
– |
– |
6,24 |
|
6,45 |
0,91 |
63,69 |
– |
– |
|
6,80 |
– |
5,51 |
– |
– |
Визначення важких металів у зрілому насінні видів роду Acer L. В умовах техногенного забруднення атмосфери і ґрунту деревні рослини акумулюють важкі метали в надлишкових кількостях, що спричиняє зміни в процесах метаболізму. Важкі метали за темпами їх надходження в біосферу і рівнем токсичності представляють найбільшу небезпеку для навколишнього середовища в порівнянні з іншими викидами промисловості [39]. Істотний інтерес становить вивчення накопичення важких металів в зрілому насіння представників роду Acer L. за умов викидів автотранспорту.
Результати дослідження щодо накопичення важких металів приведені в таблиці 3.15.
Основний шлях підвищення рівня важких металів в навколишньому середовищі – антропогенний.
Свинець відносять до високотоксичних сполук першого класу небезпечності. Він здатен значно впливати на фізіолого-біохімічні процеси рослин, інгібувати клітинний метаболізм, збільшувати токсичність інших металів, викликати мутаційні ефекти, тощо [40].
Проведенні дослідження показали, що у клена гостролистого спостерігається вміст свинцю у моніторинговій точці та контролі майже на одному рівні, відповідно – 1,90 мг/кг і 1,83 мг/кг. А у клена ясенелистого в вміст свинцю на рівні з контролем.
В антропогенних умовах останнім часом вміст кадмію в поверхневому шарі ґрунту підвищується, навіть в сільській місцевості атмосферне надходження цього елементу перевищує його виведення з ґрунту. Симптоми пошкодження кадмієм рослин – це затримка росту, пошкодження кореневої системи, некрози насіння та листя [40].
У клена гостролистого спостерігається підвищення кадмію, в порівнянні з контролем на 18%. У клена ясенелистого вміст цього елементу знаходиться майже на однаковому рівні в забрудненій зоні та чистій відповідно – 0,16мг/кг, 0,15 мг/кг
Докази незамінної ролі нікелю в метаболізмі рослин відсутні. Проте, встановлено, що при надлишку нікелю різко знижується абсорбція поживних речовин, гальмується ріст рослин і порушується метаболізм [40].
В забрудненій зоні для клена гостролистого характерне підвищення нікелю майже на 39 % в порівнянні з чистою зоною. Для клена ясенелистого характерне зменшення вмісту нікелю на 22%, відповідно для забрудненої зони вміст становить – 4,53 мг/кг, а для контролю – 5,80 мг/кг.
Цинк є одним із головних шкідливих забруднювачів не лише міських, але і сільських районів області. Цинк надходить в рослини у формі катіону, а також має великий вплив на обмін речовин.
Вміст цинку в насінні клена гостролистого в контролі становить – 37,37 мг/кг, в моніторинговій точці – 53,34 мг/кг, тобто спостерігається підвищення на 42%. Для клена ясенелистого спостерігається незначне підвищення цинку в порівнянні з контролем.
Таблиця 3. 15
Вміст важких металів в насінні різних видів роду Acer L., мг/кг
|
Назва металу
|
A. platanoides |
A. negundo |
||||
|
Вміст металу, мг/кг |
% до контролю |
Вміст металу, мг/кг |
%до контролю |
|||
|
контроль |
дослід |
контроль |
дослід |
|||
|
Свинець |
1,90 |
1,83 |
96 |
1,13 |
0,92 |
81 |
|
Кадмій |
0,22 |
0,26 |
118 |
0,16 |
0,15 |
93 |
|
Нікель |
4,37 |
6,08 |
139 |
5,80 |
4,53 |
78 |
|
Цинк |
37,37 |
53,34 |
142 |
67,72 |
70,07 |
103 |
Примітка: похибка вибірки не перевищує 5% від середніх значень
Таким чином, за умов викидів автотранспорту спостерігається підвищення важких металів в насінні клена гостролистого. А для клена ясенелистого помітні зниження вмісту металів на моніторингових ділянках. Вважаємо, що це пов’язано з тим, що насіння було попередньо очищено від зовнішньої оболонки (адже, метою дослідження було дослідити вміст металів в сім’ядолях), і можливо важкі метали акумулюються саме в ній. Тому в наступних експериментах цілеспрямовано буде проводити дослідження насіння разом з зовнішньою оболонкою.
ВИСНОВКИ
- Виявлено, негативний вплив викидів автотранспорту на масу 1000 зрілих насінин кленів. В умовах забруднення маса 1000 насінин клена гостролистого знижується на 13%, а клена ясенелистого на 10% у порівнянні з контролем.
- Встановлено, що у дозріваючому насінні дерев з чистої зони міститься більше білку, ніж із зони, забрудненої викидами автотранспорту. У порівнянні з контролем в насінинах клена гостролистого із забрудненої зони виявлено зниження рівня білку: у серпні на 16%, у жовтні на 19%. В насінні клена ясенелистого спостерігається в серпні зниження білку на 22%, у жовтні на 18%.
- Показано, суттєві зміни активності пероксидази за умов викидів автотранспорту. Так, спостерігаємо підвищення активності пероксидази в насінні клена гостролистого в серпні відповідно на 61% і 87%, в жовтні величина збільшується на 60%, ПА на 86%. Для клена ясенелистого у порівнянні з контролем спостерігається збільшення загальної активності на 65%, а питомої на 110%. У жовтні в порівнянні з серпнем активність пероксидази знижується. Проте у порівнянні з контролем також спостерігається підвищення загальної активності на 59%, питомої активності на 90%.
- Виявлено зміни в ізоферментному складі пероксидази в процесі дозрівання за дії викидів автотранспорту. Так, у дослідному насінні з’являються нові ізоформи з певним діапазоном рН 3,55; 3,60; 3,70; 4,80.
- Зареєстровано накопичення важких металів. Так, для клена гостролистого виявлено підвищення кадмію на 18 %, нікелю на 39 %, цинку на 42 %. Для клена ясенелистого накопичення важких металів в забрудненій зоні знаходиться на одному рівні з контролем.
СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ
- Новиков, В.В. Экология, окружающая среда и человек
[Текст] / В. В. Новиков. – М. : ФАИР-ПРЕСС, 2002. – 560 с. - Сухой, А.П. Древовидные растения [Текст] / А.П. Сухой. – М. : Наука, 2006. – 46 с.
- Жилін, С. Г. Сімейство кленові (Aceraceae) [Текст] / С.Г. Жилін. – М. : Просвещение, 1998. – 264 с.
- Ильин, А.М. Экология города [Текст] / А.М. Ильин. – К .: Либра, 2000. – 464 с.
- Косаківська І.В. Фізіолого-біохімічні основи адаптації рослин до стресів [Текст] / І.В. Косаківська. – К.: Сталь, 2003. – 191 с.
- Джигирей, В.С., Основи екології та охорона навколишнього природного середовища [Текст] / В.С. Джигирей, В.М. Сторожук. – Львів. Афіша, 2002. – 272 с.
- Генкель, П.А. Физиология жаро- и засухоустойчивости растений
[Текст] / П. А. Генкель. – М.: Наука, 1997. – 280 с. - Марков, Н.В. Физиология морозоустойчивости растений. [Текст] / Н.В. Марков. – М.: Наука, 1998. – 307 с.
- Бессонова, В.П. Влияние тяжелых металлов на фотосинтез растений [Текст] / В.П. Бессонова. – Днепропетровск : ДГАУ, 2006. – 208 с.
- Паршикова, Т.В. Фізіологія рослин [Текст] / Т.В.Паршикова. –Луцьк: Терен, 2010. – 23 с.
- Аксенов, С.И. Вода и ее роль в регуляции биологических процес сов [Текст] / С.И. Аксенов. – Ижевск : Институт компьютерных исследований, 2004. – 212 с.
- Кохно, Н.А. Клены Украины.[Текст] / Н.А. Кохно. – К. : Наук. думка, 1982. – 184 с.
- Денисенко, Я.В.Сучасні проблеми природничих наук [Текст] / Я.В. Денисенко. – Ніжин, 2010. – 23с.
- Коршиков, И. Взаимодействие растений с техногенно загрязненной средой. Устойчивость. Оптимизация [Текст] / И. Коршиков, В. Котов, И. Михеенко. – К. : Наукова думка,1995. – 192 с.
- Зайцева, І.О. Дослідження феноритміки деревних рослин [Текст] / І.О. Зайцева. – ДНУ, 2003. – 39 с.
- Капустян, А.В. Морфогенетична адаптивна відповідь рослин на стрес [Текст] / А.В. Капустян, О.И. Жук // Вісник Київського нац. ун-ту. Серія «Інтродукція та збереження рослинного різноманіття. – 2009. – Вип. 26. – С. 79–81.
- Ткачук, Н. П. Техноекологія [Текст] / Н. П. Ткачук. – Сімферополь: Таврія, 2000. – 650 с.
- Карпов, В. Еколого-економічні проблеми довкілля Житомирщини [Текст] / В. Карпов, С. Сіренький, В. Данилко. – Житомир, 2001. –320 с.
- Загороднюк, І. В. Індикатори біорізноманіття степових екосистем як критерій цінності природних ядер [Текст] / І.В. Загороднюк. – Луганськ: Видавництво ЛНАУ, 2009. –120 с.
- Дьякова, Т.Н. Декоративные деревья и кустарники [Текст] / Т.Н. Дьякова. – Колос 2006. – 210 с.
- Брунс, Я.Д. Каталог садових растений [Текст] / Я.Д. Брунс. 2009. –57с.
- Васильева, К.А. Эколого-биологические особенности клена в условиях техногенного загрязнения. Доклад [Текст] / К.А. Васильева. 2011. – 10 с.
- Поляков, А.К. Интродукция древесных растений в условиях техногенной среды [Текст] / А.К. Поляков. – Донецк: Ноулидж, 2009. – 268 с.
- Шматько, И. Устойчивость растений к водному и температурному стрессам [Текст] / И. Шматько, И. Григорюк, О. Шведова. – К. : Наукова думка, 1989. – 224с.
- Національний атлас України. / гол. ред. Л.Г. Руденко. – К. : ДНВП «Картографія», 2008. – 440 с.
- Шепотьев, Ф.Л. Дендрология [Текст] / Ф.Л. Шепотьев. – К .: Вища шк., 1990. – 287 с.
- Mosyakin, S. Vascular Plants of Ukraine (A Nomenclatural Checklist) [Text] / S.L. Mosyakin, M.M. Fedoronchuk – Kiev, 1999. – 345 p.
- Кохно, М.А. Дикорослі й культивовані дерева і кущі. Покритонасінні [Текст] / М.А. Кохно, М.Н. Трофименко, Л.І. Пархоменко. – К. : Фітосоціоцентр, 2005. – 716 с.
- Bradford, M.M. A rapid and sensitive method for quantitation of microgram quantities of protein ultizing the principle of protein due binding / М.М. Bradford // Anal. Biochem . – 1976. – P. 248–254
- Бояркин, А.Н. Быстрый метод определения активности пероксидазы [Текст] / А.Н. Бояркин // Биохимия. – – Т. 16, вып. 4. – 352 с.
- Сучасні методи біохімічного аналізу рослин [Текст] / Л.В. Шупранова, В.С. Більчук, Л.В. Богуславська та інші. – Д. : ДНУ, 2011. – 80 с.
- Лакин, Г.Ф. Биометрия [Текст] / Г.Ф. Лакин. – М.: Высшая школа, 1990. – 352 с.
- Голикова, М.М. Білковий обмін вегетативних органів видів роду Acer L. У річному циклі розвитку [Текст] / М.М. Голикова, І.О. Зайцева // Запорожский медицинский журнал. – 2008. – Т.2. – Вып. 2. – С. 47–
- Шупранова Л.В. Мінливість генеративного потомства деревних рослин в умовах техногенного забруднення [Текст] / Л.В. Шупранова // Сучасні проблеми фізіології та інтродукції рослин: матеріали Всеукраїнської науково-практичної конференції до 90-річчя від дня народження професора О.Ф. Михайлова. – Дн-ськ, ДНУ, 2005. – С. 63 – 64.
- Алаудинова, Е.В. Лесной журнал // Физико-химические свойства белков меристем зимуючих почек ели и пихти [Текст] / Е.В. Алаудинова. – 2003. – 15 с.
- Алаудинова, Е.В. Белки цитоплазмы меристемы почек ели: динамика аминокислотного состава. Химия растительного сырья [Текст] / Е.В. Алаудинова. – 2002. – 25 с.
- Конарев, В.Г. Белки растений как генетические маркеры [Текст] / В. Г. Конарев. – М. : Колос. – 1983. – 320 с.
- Ветчинникова, Л.В. Морфо-физиологические и биохимические особенности различных видов и разновидностей березы семенного и вегетативного происхождения в условиях Восточной Фенноскандии: автореф. дис. д. б. н. М., 2002. – 24 с.
- Бессонова, В.П. Влияние тяжелых металлов на фотосинтез растений: Монография [Текст] / В.П. Бессонова. – Д: ДГАУ. – 2006. – 208 с.
- Грицан, Н.П. Оценка состояния и уровня загрязнения тяжелыми металлами фитоценозов города Днепропетровска [Текст] / Н.П. Грицан. – Д., 1992. – 66 с.
Редакція може не поділяти думку авторів і не несе відповідальність за достовірність інформації. Будь-який передрук матеріалів з сайту може здійснюватись лише при наявності активного гіперпосилання на e-kolosok.org, а також на сам матеріал!
Поширити


Схожі публікації

КОЛОСОК ДЛЯ ЗСУ 9-11 клас
Запрошуємо до участі у конкурсі усіх великих і малих ))), і не думайте що якщо Ви великі, то вам буде легко )

КОЛОСОК ДЛЯ ЗСУ 7-8 клас
Запрошуємо до участі у конкурсі усіх великих і малих ))), і не думайте що якщо Ви великі, то вам буде легко )

КОЛОСОК ДЛЯ ЗСУ 5-6 клас
Запрошуємо до участі у конкурсі усіх великих і малих ))), і не думайте що якщо Ви великі, то вам буде легко )